Contents |
Introduction
This fact sheet has been developed to support the implementation of the Natural Resources Conservation Service Feed Management 592 Practice Standard. The Feed Management 592 Practice Standard was adopted by NRCS in 2003 as another tool to assist with addressing resource concerns on livestock and poultry operations. Feed management can assist with reducing the import of nutrients to the farm and reduce the excretion of nutrients in manure.
The Natural Resources Conservation Service has adopted a practice standard called Feed Management (592) and is defined as “managing the quantity of available nutrients fed to livestock and poultry for their intended purpose”. The national version of the practice standard can be found in a companion fact sheet entitled “An Introduction to Natural Resources Feed Management Practice Standard 592”. Please check in your own state for a state-specific version of the standard.
Nitrogen (N) is the building block of proteins in feeds and forages. Protein is typically the most expensive component of the purchased feeds used in dairy rations. Nitrogen is also receiving more attention as a component of nutrient management plans on dairy farms and potential ammonia emissions.
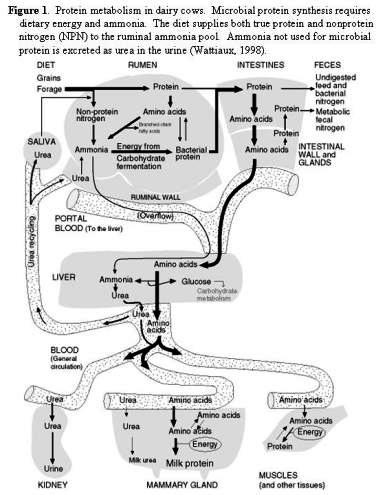
Understanding how N is used in dairy cattle is important in improving both profitability and decreasing excretion from the cow into the environment. It is important to remember that dairy cows do not have a protein requirement. They really need amino acids available in the small intestine to support tissue growth and milk production. Basically, N utilization in dairy cattle is composed of two components. The first is providing an adequate supply of N and carbohydrates in the rumen to support the growth of rumen microorganisms and the production of microbial crude protein (MCP). The second part of the system is the utilization of amino acids in the small intestine to provide for the needs of the cow.
Please check this link first if you are interested in organic or specialty dairy production
Definitions
N = nitrogen; CP = crude protein; NPN =nonprotein nitrogen; TDN = total digestible nutrients; MP = metabolizable protein; MCP = microbial crude protein; SP = soluble protein; RDP = rumen degradable protein; RUP = rumen undegradable protein; NRC = National Research Council
Feed Nitrogen Fractions
Even though all feeds contain N, there is variation in the quantity of N in each feed and it’s availability and utilization in the dairy cow. Forage testing laboratories determine the quantity of N in the sample and multiply this value by 6.25 to obtain the crude protein (CP) value printed on the analysis report. This calculation assumes that feeds contain 16% N on a dry matter (DM) basis. An example calculation is:
Alfalfa silage = 3% N * 6.25 = 18.75% CP (both on a DM basis)
The challenge is that feeds could have the same CP value, but have a different feeding value to the dairy cow. Consider the following examples:
Alfalfa hay, alfalfa silage and alfalfa pasture – All 20% CP.
Raw and roasted soybeans – Both with 40% CP
Even though these feeds have the same CP level, we would not expect the same level of N utilization and milk production. If we are feeding 4 pounds/cow/day of raw soybeans to a dairy cow producing 80 pounds of milk, replacing these with 4 pounds of roasted soybeans would increase predicted milk production on a protein basis by 2-3 lbs. What is the reason for this?
One reason is that there are a number of N compounds found in feeds. This means that we need to better define the types of N compounds present in feeds. A simple to start is to classify feed N as either true protein or NPN. These can be defined as:
True protein = The N in feeds found in complex and linked structures as amino acid combinations. Examples are: albumins, globulins and amino acids. These feeds will vary in both the rate and extent of degradation that occurs in the rumen.
NPN = This is the N in simple compounds such as ammonia or urea (not as amino acids). These are considered to be rapidly available in the rumen.
The above breakdown is a start, but the true protein component needs to be better defined for use in ration formulation or evaluation programs. This is most commonly done in the following manner:
RDP = that portion of the total N intake that is degraded in the rumen. The NPN fraction is included in RDP.
RUP = that portion of the total N that is not degraded in the rumen and passes intact to the small intestine. There is a portion of the RUP fraction that is not available or digested in the small intestine and passes out in the feces. This is fraction C in the system described by Van Soest (1994).
Ruminal N Metabolism
A portion of the feed N that enters the rumen will be degraded to compounds such as peptides, amino acids or ammonia. The primary mechanism for this breakdown in the rumen is microbial proteolysis. The solubility, structure, and particle size of the feed will all influence the amount of degradation that takes place. There will always be a portion of the feed N that enters the rumen that is not degraded (RUP).
All RDP does not breakdown and be converted to ammonia at the same rate. Van Soest (1994) provided an overview of a system to define N sub-fractions that would permit better characterization of feed N availability and use in the dairy cow. This system includes the following fractions:
A – This is mainly NPN, amino acids, and peptides that are “instantly” available in the rumen.
B1 – This fraction has a fast rate of degradation in the rumen.
B2 – This fraction has a variable rate of degradation in the rumen.
B3 – This fraction has a slow rate of degradation in the rumen.
The use of this approach assists in doing a better job of describing N utilization in the rumen and improving the efficiency of feed N use. The use of this approach does require additional feed analysis data and computer formulation programs designed to utilize this information.
Microbial Protein
Microbial protein (MCP) is produced in the rumen by the rumen microorganisms. The key factors that determine the quantity of MCP synthesized is the quantity of ammonia available in the rumen and the supply of fermentable carbohydrates to provide an energy source. The availability of peptides may also stimulate the production of MCP by some rumen microorganisms. The NRC (2001) predicts MCP production as 13% of the discounted TDN (total digestible nutrients) available in the rumen.
Microbial protein can provide 50 – 80% of the amino acids required in the intestine by the dairy cow. Optimizing MCP production helps in increasing the efficiency of N use in the cow and controlling feed costs.
The benefits of MCP are related to:
- MCP averages about 10% N (60-65% CP).
- MCP is a good source of RUP.
- MCP has a high digestibility in the intestine.
- The amino acid profile of MCP is fairly constant.
- MCP has an excellent ratio of lysine to methionine.
Protein Systems
There are 2 systems used to evaluate and balance rations for dairy cows on a protein basis. These are the CP (crude protein) and MP (metabolizable) protein systems. The CP system has been the most commonly used system.
The CP system is easy to use and has tabular feed composition and animal requirement information. This system assumes that all N in different feeds is similar in use and value to the cow. The Dairy NRC (2001) indicated that CP was a poor predictor of milk production. Nutritionists have modified the CP system to better meet their needs. They have added SP, RDP and RUP as additional factors to consider when using CP as the base for formulating dairy rations on a protein basis.
The Dairy NRC (2001) has suggested moving to a MP system to better define and refine protein formulation and utilization. This system fits with the biology of the cow. The challenge is that this system is not tabular and requires the use of computer programs to calculate both MP requirements and the MP supplied by feeds and MCP. The industry is changing to an MP approach. This system should provide an opportunity to improve the efficiency of protein use in dairy cattle. The use of this system will also decrease N excretion to the environment and lower potential ammonia emissions.
Total N Use in Dairy Cows
It is important to realize that the dairy cow is a dynamic rather than static system. This means that the actual value of a feed N source will vary depending on a number of factors. These include:
- The proportion of the total N intake used in the rumen versus the small intestine.
- The length of time the feed remains in the rumen (rate of passage).
- The rate at which the feed is degraded in the rumen (rate of digestion).
- The amino acid profile of the RUP fraction.
- The digestibility of the RUP and MCP fractions in the small intestine.
This situation is similar to the energy value of feeds that occurs due to differences in dry matter intake (DMI) and rate of passage. Dairy cows with higher levels of DMI have a higher rate of passage and lower feed energy values. This is the reason for discounting feed energy values based on level of DMI and milk production (NRC, 2001).
The NRC (2001) computer model was used to determine the RDP and RUP for soybean meal in a ration for dairy cows. The base ration was for a cow producing 80 pounds of milk and contained 5 pounds of DM from soybean meal. This ration was then evaluated for cows producing 60, 100 or 120 lbs. of milk. The ration ingredients were all kept in the same proportion, but total ration DMI was adjusted using the NRC program predicted intakes. This would be similar to cows fed a 1-group TMR. The RDP and RUP values for soybean meal in this ration were:
| Milk, lbs/day | RDP, % of CP | RUP, % of CP |
| 60 | 60 | 40 |
| 80 | 59 | 41 |
| 100 | 56 | 44 |
| 120 | 54 | 46 |
The reason for the higher RUP value in higher producing cows is the decreased amount of time the soybean meal stays in the rumen. Thus, there is less time for N degradation and proteolysis to take place. This example also indicates the challenge with using tabular values to describe the RDP and RUP fractions in feeds. This is the reason that computer programs that can integrate DMI, rate of passage and rate of digestion are needed as we continue to refine formulation and evaluation approaches.
Summary
Nitrogen is the most expensive component of purchased feed costs on most dairy farms. Ration programs that incorporate the concepts of feed fractions and variable feed contributions to the animal provide an opportunity to fine tune nutrition and improve the efficiency of nutrient use. This will also lower nutrient excretion to the environment and usually improves income over feed cost.
References
NRC, 2001. National Research Council. Nutrient Requirements of Dairy Cattle. 7th rev. ed. National Academy of Science, Washington, DC.
Van Soest, P.J. 1994. Nutritional ecology of the ruminant. Cornell University Press, Ithaca, NY.
“Extension programs and policies are consistent with federal and state laws and regulations on nondiscrimination regarding race, sex, religion, age, color, creed, national or ethnic origin; physical, mental or sensory disability; marital status, sexual orientation, or status as a Vietnam-era or disabled veteran. Evidence of noncompliance may be reported through your local Extension office.”
Disclaimer
This fact sheet reflects the best available information on the topic as of the publication date. Date 4-12-2007
This Feed Management Education Project was funded by the USDA NRCS CIG program. Additional information can be found at Feed Management Publications.
![]()
This project is affiliated with the Livestock and Poultry Environmental Learning Center.
![]()
Project Information
Detailed information about training and certification in Feed Management can be obtained from Joe Harrison, Project Leader, jhharrison@wsu.edu, or Becca White, Project Manager, rawhite@wsu.edu.
Author Information
L.E. Chase
Cornell University
lec7@cornell.edu
Reviewer Information
Mike Hutjens – University of Illinois
Floyd Hoisington – Consulting Nutritionist
Partners